Германское издательство LAP LAMBERT Academic Publishing (LAP LAMBERT Academic Publishing is a trademark of: OmniScriptum GmbH & Co. KG) предлагает книгу на русском языке "Исторические исследования" (ISBN: 978-3-659-63167-2, цена - 89.90 ЕВРО), автор - Тихомиров Андрей Евгеньевич, аннотация на обложке "Современные археологические исследования доказывают, что родиной индоевропейцев является район Южного Урала, где они сформировались в качестве единой языковой группы. Аирйанэм-Ваэджа (авест. «арийский простор») — мифическая прародина древних иранцев, ариев; согласно Видевдату (Вендидаду, первой книге Авесты, сборника священных книг древнеиранской религии, своего рода иранское продолжение Вед), именно с неё начинается список из 16 «наилучших» «местностей и областей», созданных Ахура Маздой для человечества. Эта страна описывается как бескрайняя равнина, через которую протекает прекрасная река Даитйя (Вахви-Датия) - Урал". Заказать книгу:https://www.morebooks.shop
Статьи Тихомирова А.Е.
ПЛАНЕТА ОБЕЗЬЯН
4 сентября 2013 г.
Земля, хотим мы того или нет, захвачена обезьянами, называющие себя первыми (примат от лат. primatus – первое место, старшинство) и как ни крути, но так называемые люди разумные (Homo sapiens) это всего лишь один из ныне живущих 200 видов – от лемуров до человека отряда приматов или обезьян. Ну, а какие обезьяны, в той или иной степени многим известно: крикливые, шумные, с инстинктом стадности, с резко выраженной иерархической системой доминирования-подчинения, у многих видов обезьян господствует матриархат. Эмоции, гримасничанье, паясничанье и т.д. у обезьян зачастую превалируют над разумом. Всё это имеет место и у человека – первой обезьяны планеты.
Находки останков различных человекоподобных существ, таких, как питекантропы, кроманьонцы и неандертальцы, которых современная наука считает предками человека, не редкость. Подобные находки периодически делаются археологами в очень древних слоях - свыше 30, 50 и даже 100 тысяч лет.
Современная наука считает, что создание человека связано с генетическими мутациями, древними вирусами. Различные вирусы, встраиваясь в геном древних гоминид (человекообразных существ) привели к формированию многообразных человекоподобных существ. Из них в конечном итоге, в ходе бесконечной борьбы за существование победил современный вид – homo sapiens. В то же время существовали и другие виды человекообразных существ – гоминид, исследования открытых недавно в Кении (восточная Африка) могильников свидетельствует о том, что homo habilis (человек умелый) и homo erectus (человек прямоходящий) жили бок о бок. Они могли спариваться и, по-видимому, из этого промискуитета (ничем не ограниченные половые отношения) и произошел современный вид - homo sapiens. А все многообразие человекоподобных существ, современные расы – это итог бесконечной борьбы за существование, когда человекоподобные существа вступали в половые контакты с близкими видами и не только, например, известны различные формы половых отношений - скотоложство, некрофилия, гомосексуализм и др.
Французские ученые из Университета Руана обнаружили в 2009 году новый штамм вируса иммунодефицита человека (ВИЧ). По их данным, он передается человеку от горилл. Это первый зафиксированный случай заражения ВИЧ от приматов за исключением шимпанзе - три известные до сих пор вида вируса иммунодефицита перешли к человеку именно от этих приматов. Вирус был обнаружен у женщины французского происхождения, проживающей на территории Камеруна. Она не употребляла в пищу обезьянье мясо, поэтому ученые полагают, что заражение произошло половым путем от другого человека.
Ученые из российского Института молекулярной генетики РАН утверждают, что человека из древней гоминиды сделали древние вирусы. «Гены, как уже можно утверждать, у человека и обезьяны сходны, они выделяют обычные белки, но вирусы меняют их соотношение, распределение и затраченное на это время», пояснил заместитель директора института Вячеслав Тарантул. Почти одновременно с россиянами озвучили свою теорию превращения обезьяны в Homo sapiens и американцы. Как заявили биологи и специалисты по пластической хирургии из Университета штата Пенсильвании, им удалось обнаружить единственную генетическую мутацию, благодаря которой древнейшие предки человека начали отличаться от своих обезьяноподобных предшественников. Фактически, речь идет о моменте рождения человека на планете Земля. Эта мутация, уверяют ученые, произошла примерно 2,4 млн. лет назад в гене, который контролирует развитие челюстных мышц и у современного человека называется MYH16. В результате этого, казалось бы, небольшого изменения в геноме челюстные мышцы наших предков стали ослабевать, что заставило несчастных менять свои привычки, меню, искать первые орудия труда. Им пришлось приспосабливаться к новым особенностям организма, учиться готовить еду, как-то ее обрабатывать. Именно изменение челюстных мышц, которые, став менее мощными, оказывали меньшее давление на череп, обеспечило иное его строение, позволив развиться куда более крупному головному мозгу, этот мощный «компьютер» обслуживается хорошо развитыми органами чувств. В итоге обезьяноподобное существо взяло в лапы палку, и решило больше ее из них не выпускать. Этим возможно объяснить и то, что проводящие пути, идущие от головного мозга к спинному у человека, перекрещиваются, так что правое полушарие управляет левой половиной тела, а левое – правой. А вот остальные обезьяны такой мутации были лишены и до сих пор обладают мощными челюстями и слабым, относительно человека, мозгом. У человеческих детенышей швы, соединяющие кости черепа, могут раздвигаться при росте и имеются роднички – участки мозга, не прикрытые костью. С возрастом роднички зарастают, и по достижении мозгом нормальной величины размеры черепа уже не увеличиваются. Однако рост мозга продолжается и у многих современных детей роднички долго не зарастают. Библейский рассказ о проклятии Евы «в болезни будешь рожать детей» восходит к древнему «божественному» объяснению, почему женщины рожают с болью. Обезьяны и человек остались всеядными, не специализируясь к какой-либо пище, поэтому зубная система у них близка к исходному типу, сформировавшемуся еще у древних зверозубых ящеров - пресмыкающихся. Человек обладает цветным бинокулярным зрением, как у обезьян. Человекообразные обезьяны жили в условиях достаточно постоянного климата и не имели четко выраженного сезона размножения, поэтому у женщин развился менструальный цикл, длящийся 28 дней, то есть 13 раз в год возможно зачатие, цикл находится под контролем эндокринной системы.
Мутация произошла только у одной популяции древних гоминид, остальные особи из этой популяции, скорее всего, погибли. Гибель, возможно, произошла по причине какой-либо болезни, инфекции, облучения, заражения и т.д., что и привело к значительному сокращению популяции древних гоминид, оставшиеся (условные Адам и Ева) смогли выжить ослабленными и дать потомство с генномодифицированными мускулами скул (скуловая дуга – наследие зверозубых ящеров), эти гены оказались доминантными, спариваясь с близкими сородичами из других популяций, эта мутация закрепилась и развилась дальше у человеческих предков.
Ранее считалось, что загадка в тех генах, которые отличны у человека и у обезьян, в 2006 году даже был открыт участок генома, изменившийся у человека сильнее всего, однако это не внесло ясности. Возможно, что отличия между обезьяной и человеком кроются не только в разных генах, но и в одинаковых. Ральф Хэйгуд и его коллеги из университета Дюка смогли доказать верность этой догадки. Ученые проанализировали 6280 генов, которые присутствуют у человека, шимпанзе и макаки. Оказалось, что у 575 человеческих генов, многие из которых были связаны с работой мозга и, что более удивительно, питанием, промоутеры имеют сильные отличия от своих аналогов у обезьян. Промоутеры - это молчащие гены, которые при благоприятных условиях выполняют регулятивную функцию, то есть руководят работой других генов. Промоутеры при необходимости вырабатывают белки, которые в свою очередь влияют на работу других генов. Это ученые посчитали признаком положительного эволюционного отбора, который помог нам стать теми, кто мы есть. Интересно, что участки ДНК, которые сильнее всего отличают обезьяну от человека, связаны с работой мозга и с питанием. Более 100 генов из 575 отличных участков связаны с метаболизмом углеводов, а особенно глюкозы. Хейвуд полагает, что некогда энергию для развития мозга человек получил благодаря изменениям в рационе — переходу от фруктов к клубням и корням.
В 2007 году ученые Кембриджского университета выяснили, что знаменитое выражение «ВСЕ ЛЮДИ – БРАТЬЯ» является не просто метафорой, согласно последним научным исследованиям, все население Земли происходит от одного маленького африканского племени, которое появилось в результате генетической мутации около 200 тысяч лет назад в районах Центральной и Южной Африки, а спустя 150 тысяч лет эти первые люди начали мигрировать на другие континенты. Исследования велись в двух направлениях, ученые изучали генетический код различных народов Земли и физические особенности 6 тысяч черепов древних людей, обнаруженных в различных частях планеты. Одновременно установлено, что сосуществовали и другие «люди» - представители других ветвей человечества. Научно доказано, что неандертальцы и homo erectus иммигрировали из Африки намного раньше наших прямых предков.
Группа американских ученых ранее установила, что мозг человека продолжает эволюционировать, сравнивая генотип современного человека и предка, жившего 37 тысяч лет назад, исследователи обнаружили значительное расхождение в строении двух генов, отвечающих предположительно за объем головного мозга, что связано с мутациями. Основные изменения в первом гене (microcephalin) связаны с крупными культурными явлениями, такими, как музыка, искусство, письменная речь, а также требующие сложных навыков ремесла. Эта мутация произошла примерно 50 тысяч лет назад и в настоящее время присутствует у 70% жителей Земли.
Другая мутация произошла, по эволюционным меркам, совсем недавно – 5800 лет назад, такой ген присутствует примерно у 30% ныне живущих людей, вариант второго гена (ASPM) появился как следствие развития сельского хозяйства, возникновения городов и первых дошедших до нас образцов письменности. Кстати, существуют и гены бродяжничества, лазания по деревьям, жестокости и другие. Руководитель исследования доктор Брюс Лан говорит о том, что эти два гена объясняют тенденцию постепенного увеличения объема головного мозга и усложнения его строения, которое наблюдается у людей, и этот процесс будет продолжаться вечно. Ведь, по словам Карла Маркса, бытие определяют сознание, и если создаются соответствующие условия выживания, окружающей среды, то в той или иной мере мозг человека воспроизводит древние гены. Например, у бродяг (лиц без определенного места жительства) «просыпаются» гены бродяжничества, в криминальной среде – ген наживы и алчности, в музыкальной среде - гены музыкальной одаренности и т.д.
На вопрос о том, какие виды обезьян относятся к группе человекообразных, многие люди, не задумываясь, отвечают: шимпанзе, горилла, орангутанг. Те, кто более сведущ в зоологии, называют еще гиббона. Действительно, эту обитающую в Юго-Восточной Азии обезьяну также включают в надсемейство человекоподобных (Hominoidea). Хотя, если судить только по внешнему виду, на человека гиббон не очень-то и похож. А вот о существовании нашего куда более близкого родственника, бонобо, или карликового шимпанзе, мало кто знает. И это притом, что набор генов бонобо совпадает с набором генов человека на 98%! Международная группа экспертов в 2009 году обнаружила самый древний скелет человекообразного существа, возраст которого 4,4 миллиона лет. «Это не шимпанзе. Это не человек. Это доказывает, какими мы были», - заявил сотрудник Университета в Беркли.
Полностью расшифрован в 2012 году геном гориллы, которая оказалась более близким родственником человека, чем шимпанзе. Генетики объясняют это эффектом быстрого видового разделения гоминид, случившегося 10 млн. лет назад. Примерно 15% генов гориллы ближе к их человеческим аналогам, чем к тем, которые содержатся в ДНК шимпанзе, говорится в статье, опубликованной в журнале Nature.
Американские генетики рассчитали в этом году возраст Адама и Евы. Учёные установили, что Y-хромосомный Адам существовал на Земле 120-156 тысяч лет назад, что пересекается с диапазоном существования митохондриальной Евы. По словам доктора Карлоса Бустаманте из Стэнфорда, благодаря работе его команды устранены расхождения во времени существования генетических предков людей, согласно которым более молодой Адам мог никогда не встретить свою Еву, что неприятно, даже по меркам каменного века. «В течение длительного времени это была головоломка в генетике человека, ― рассказывает профессор Бустаманте. ― Предыдущие исследования показали, что последний общий предок мужчин жил 50-115 тысяч лет назад, что значительно позже женского предка. Теперь же мы устранили расхождения, и наш Y-хромосомный Адам даже немного старше». Когда учёные используют термины «последний общий предок», или «Y-хромосомный Адам» речь идёт не о конкретном человеке, а скорее, о генетически однородной общности людей. Y-хромосомы и митохондрии именно таких абстрактных популяций унаследовало современное человечество. Чтобы проследить генетических предков мужчин и женщин, учёные выбирают последовательности ДНК Y-хромосом и митохондрий из-за устойчивости их генетического материала во времени, а ещё из-за особенности их наследования: от отца к сыну и от матери к дочери соответственно.
Y-хромосомный Адам - понятие археогенетики, обозначающее наиболее близкого общего предка всех ныне живущих мужчин. Он назван в честь библейского Адама, но не был единственным мужчиной своего времени. Предшественники и современники Y-хромосомного Адама имели другую Y-хромосому, однако их род по мужской линии по какой-то причине прервался. Поэтому все последующие поколения мужчин являются носителями гаплогруппы Y-хромосомного Адама. Митохондриальная Ева – сходное молекулярно-биологическое понятие. Это имя, данное женщине, которая была самым древним общим предком современных женщин. Митохондриальная ДНК наследуется только по материнской линии: если у женщины нет ни одной дочери, ее митохондриальная ДНК будет передана потомкам не дальше ее сына. Определение того, когда жили Y-хромосомный Адам и митохондриальная Ева, является очень сложной задачей. Для этого ученые используют метод молекулярных часов: зная количество оснований в хромосоме и частоту их мутаций, можно оценить, сколько времени прошло с момента возникновения хромосомы. В процессе изучения этого вопроса данные о датах жизни Адама и Евы сильно менялись. Так, в 2000 году было сделано предположение, что Y-хромосомный Адам жил около 59 тысяч лет назад, последние исследования показали более древнюю дату - 115 тысяч лет назад. Тем не менее, ученые сходились во мнении, что митохондриальная Ева существовала намного раньше Адама - примерно 150-240 тысяч лет назад, с максимальной вероятностью этого значения 180-200 тысяч лет назад. Группа ученых из США и Франции, возглавляемая Дэвидом Позником из Стэндфордского университета, утверждает: общие предки современных людей по мужской и женской линии существовали примерно в одно время. С результатами работы группы можно ознакомиться в авторитетном научном журнале Science. Ученые изучили Y-хромосомы 69 мужчин, относящихся к 9 различным популяциям (в соответствии с данными проекта по разнообразию геномов человека (the Human Genome Diversity Project) , который осуществляется Стэндфордским университетом). Мужчины относились к следующие народам: бушмены (Намибия), пигмеи мбути (Демократическая Республика Конго), пигмеи бака и нджембе (Габон), берберы (Алжир), пуштуны (Пакистан), якуты (Сибирь), майя (Мексика), камбоджийцы. В результате секвенирования ДНК (определения аминокислотной и нуклеотидной последовательности) и построения «дерева», отражающего ответвления новых групп от основной ветви, исследователи получили новые данные: они предполагают, что Y-хромосомный Адам жил приблизительно 120-156 тысяч лет назад, причем пик вероятности приходится на период 138 тысяч лет назад. Группа Дэвида Позника провела аналогичные исследования митохондриальных ДНК женщин из семи разных популяций и установила, что митохондриальная Ева жила 99-148 тысяч лет назад (максимальная вероятность - 124 тысячи лет назад). Ученые предполагают, что большой разброс данных, полученных в ходе ранней работы, обусловлен материалом для исследований: это были короткие перегруппированные сегменты ДНК, которые не могли дать достаточно точной информации. Теперь же, изучив эталонную последовательность митохондрий, можно достаточно аргументированно утверждать: Адам и Ева, наши генетические прародители, существовали практически одновременно.
На правах рукописи УСТИНОВА Галина Михайловна
ХАРАКТЕРИСТИКА МАКРОФИТНО-БАКТЕРИАЛЬНЫХ ВЗАИМООТНОШЕНИЙ В ВОДНЫХ БИОЦЕНОЗАХ
03.00.16 -«Экология»
АВТОРЕФЕРАТ
диссертации на соискание ученой степени кандидата биологических наук
Оренбург – 2004
Работа выполнена в Государственном образовательном учреждении высшего профессионального образования «Оренбургской государственной медицинской академии Министерства здравоохранения Российской Федерации».
Научный руководитель: Заслуженный работник высшей школы, доктор биологических наук, профессор Соловых Г.Н.
Официальные оппоненты: Доктор медицинских наук Гриценко В.А.
Доктор биологических наук, профессор Рябинина З.Н.
Ведущая организация: Институт экологии и генетики микроорганизмов УрО РАН
С диссертацией можно ознакомиться в библиотеке Оренбургской государственной медицинской академии.
Ученый секретарь диссертационного совета, доктор медицинских наук, профессор Немцева Н.В.
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы.
Практически все живые организмы на планете существуют в тесной связи друг с другом, находясь в симбиотических отношениях (Бухарин О.В., 1999). Показано, что своеобразие биоценозов, формирующихся в различных биотопах, обусловлено совокупным действием комплекса абиотических факторов (Шварц С.С., 1980; Одум Ю., 1986), а так же биологическими особенностями членов сообщества, определяющими характер взаимоотношений между ними (Бонда-ренко В.М. и др., 1997; Воронов А.В., 2000). Жизнь водоема, его биоценозов, характеризуется эволюционно сложившимися взаимоотношениями между макро- и микроорганизмами в экосистеме (Константинов А.С., 1967; Сергеев В.П., 2000). Хотя водные макрофиты и бактерии являются неотъемлемой частью гидробиоценозов, механизмы формирования макрофитно-бактериальных ассоциаций в настоящее время наименее изучены. Литературные данные по изучению их взаимоотношений обычно ограничиваются трофическими связями (Сомов Г.П. и др., 1988; Аксенов М.Ю. и др., 1994; Горобец О.Б., 2001).
Многочисленными исследованиями показано, что одним из экологически значимых факторов, принимающих участие в формировании гидробиоценозов, является лизоцим (Багнюк В.М. и др., 1980; 1983). Обосновано существование в пресных водоемах особой функциональной системы «лизоцим гидробионтов - антилизоцим бактерий» (Соловых Г.П., 1995), способствующей формированию простых и сложных сетевых связей между гидробионтами. Г.Н. Соловых и И.В. Карнаухова (1999) выделили лизоцим пресноводного двустворчатого моллюска Unio pictorит и показали специфические его действия на водные микроорганизмы. Показана роль лизоцима как одного из биологических факторов гидробионтов, участвующих в процессах самоочищения водоемов от патогенной и условно патогенной микрофлоры (Нефедова Е.М., 2001). Однако, роль функциональных связей типа «лизоцим-антилизоцим» в формировании макрофитно-бактериальных ассоциаций остаются изучены недостаточно. Мало сведений о таксономическом спектре микроорганизмов -ассоциантов макрофитов. Недостаточно исследованы биологические свойства, характерные для бактерий и макрофитов в условиях природных сообществ, в частности нет данных о распространенности и выраженности лизоцимного и антилизоцим ного признаков в макрофитно-бактериальных сообществах водоемов. До сих пор недостаточно изучен вопрос о роли макрофитов в формировании различных сообществ гидробионтов и бактериопланктона в том числе.
Как фототрофные организмы макрофиты обладают многообразием синтетических процессов, конечными продуктами которых являются биологически активные вещества (Гудвин Т. и др., 1986), в частности флавоноиды. Микроорганизмы контактируют в среде своего обитания с флавоноидами, которые могут быть пищевыми субстратами для некоторых бактерий (Winter J., 1989; Filial B.V.S., 2002), или выполнять определенную роль в симбиотических взаимоотношениях с микроорганизмами (Pandya S., 1999) и участвовать в защите растений от инфекций (Padmavati M., 1997). В этой связи актуальной является оценка роли флавоноидов в формировании макрофитно-бактериальных ассоциаций.
Таким образом, важное значение макрофитно-бактериальных ассоциаций в экологии водоемов, а также отсутствие сведений о механизмах их формирования послужили основой для проведения данного исследования. Цель исследования. Изучение биологического разнообразия макрофитно-бактериальных сообществ в водных биоценозах и оценка характера эколого-функциональных связей, способствующих формированию ассоциаций макрофитов с бактериями. Задачи исследования:
1. Определить биологическое разнообразие природных макрофитно-бактериальных сообществ в водных экосистемах различных типов (лотические и лентические).
2. Охарактеризовать биологические свойства бактериальной флоры (их морфологическое разнообразие, лизоцимную и антилизоцимную активность) и макрофитов (уровень лизоцимной активности, содержание флавоноидов и аскорбиновой кислоты) в природных водоемах.
3. Изучить роль флавоноидов и аскорбиновой кислоты макрофитов в образовании макрофитно-бактериальных ассоциаций водных биоценозов.
4. Оценить значение системы «лизоцим макрофитов-антилизоцим бактерий» в формировании эколого-функциональных связей в макрофитно-бактериальных сообществах водных экосистем. Научная новизна. Проведена оценка биологического разнообразия макрофитно-бактериальных сообществ водных экосистем на территории Оренбургского района, выявлены доминирующие виды и эколого-морфологические особенности макрофитов и микрофлоры в лотических и лентических экосистемах. Дана оценка содержания общего количества флавоноидов (в том числе, рутина) и аскорбиновой кислоты у водных макрофитов. Показано, что их содержание варьирует у разных видов и зависит от периода вегетации (бутонизация и цветение). Выявлена прямая корреляция численности бактерий-ассоциантов с содержанием в макрофитах общего количества флавоноидов и аскорбиновой кислоты, отражающая участие данных биологически активных веществ в формировании макрофитно-бактериальных ассоциаций.
Обнаружена лизоцимная активность в экстрактах тканей и показана вариабельность ее уровня у доминантных видов водных макрофитов. Среди бактерий-ассоциантов макрофитов отмечена распространенность антилизоцимной активности, однако не было обнаружено бактерий-ассоциантов с лизоцимной активностью. Выявлена прямая корреляционная зависимость между уровнем лизоцимной активности макрофитов и численностью антилизоцимактивных бактерий, что свидетельствует о роли системы «лизоцим макрофитов - антилизоцим бактерий» в формировании эколого-функциональных связей в макрофитно-бактериальных сообществах водных биоценозов. Научно-практическая значимость. Полученные данные расширяют теоретические представления о биоразнообразии и эколого-функциональных взаимоотношениях макрофитов и бактерий в лотических и лентических экосистемах. В результате проведенных исследований обосновано участие биологически активных веществ (флавоноидов, аскорбиновой кислоты) макрофитов и роли системы «лизоцим макрофитов антилизоцим бактерий» в формировании макрофитно-бактериальных ассоциаций в водных биоценозах. Разработан «Способ биологического обеззараживания непроточных водоемов от патогенной и условно-патогенной микрофлоры» (патент РФ на изобретение № 2193531 от 27 ноября 2002 года), основанный на интродукции в пруды биологической очистки гидробионтов, обладающих лизоцимной активностью. Внедрение результатов исследования в практику. Результаты исследования внедрены и используются в Центрах Госсанэпиднадзора Оренбургской области и г. Оренбурга (Акты внедрения от 25.05.04, 27.05.04); в научно-производственной фирме «Экобиос» (Акт внедрения от 21.05.04); на кафедре физиологии растений ГОУ ВПО ОГПУ (Акт внедрения от 12.03.04); на кафедре биоэкологи и зоологии ОГАУ (Акт внедрения от 12.03.04). Апробация работы. Основные положения диссертационной работы доложены и обсуждены на II международной конференции «Биоразнообразие и биоресурсы Урала и сопредельных территорий», г. Оренбург, 2002; на межрегиональной конференции молодых ученых «Современные проблемы экологии, микробиологии и иммунологии», г. Пермь, 2002; на Всероссийской научно-практической конференции, посвященной 80-летию создания госсанэпидслужбы России, Москва, 2002; на региональной научно-практической конференции молодых ученых и специалистов, г. Оренбург, 2003; на межрегиональной конференции биохимиков Урала, Западной Сибири и Поволжья «Биохимия: от исследования молекулярных механизмов - до внедрения в клиническую практику и производство», г. Оренбург. 2003; на международной научно-практической конференции «Актуальные проблемы ветеринарной медицины и биологии», г. Оренбург, 2003; на региональной научно-практической конференции молодых ученых и специалистов, г. Оренбург, 2004; на II Всероссийской конференции «Биоразнообразие, экология, охрана», г. Борок, 2004. Положения, выносимые на защиту:
1. Биоценозы лотических и лентических водоемов характеризуются биологическим разнообразием макрофитно-бактериальных сообществ, уровень которого зависит от типа экосистемы.
2. Биологически активные вещества макрофитов - флавоноиды и аскорбиновая кислота и система «лизоцим макрофитов - антилизоцим бактерий» участвуют в формировании макрофитно-бактериальных ассоциаций водных биоценозов. Объем и структура диссертационной работы.
Диссертация изложена на 149 страницах машинописного текста, состоит из введения, 5 глав, заключения и выводов. Указатель литературы содержит 126 отечественных и 21 зарубежных источника литературы. Текст иллюстрирован 21 таблицами и 29 рисунками. Публикации. По теме диссертации опубликовано 9 печатных работ, 1 из них в центральном журнале; получен патент РФ на изобретение № 2193531 от 27 ноября 2002 года. Связь работы с научными программами. Диссертационное исследование «Характеристика макрофитно-бактериальных взаимоотношений в водных биоцендзах» проводится в рамках темы открытого плана НИР ОрГМА МЗ РФ (№ гос.регистрации 01.20.02 15158). СОДЕРЖАНИЕ РАБОТЫ
Материалы и методы. Работа выполнена на р. Урал и двух озерах в районе г. Оренбурга (оз. Коровье и оз. Микутка) в период с 2001 по 2003 годы, а также в условиях лабораторного эксперимента. Основным объектом исследования явились макрофиты и микрофлора воды исследуемых водоемов.
Химический анализ воды проводился согласно общепринятым в гидрохимии методам, описанным в руководстве О.А. Алекина (1973).
Полевые геоботанические исследования проводились в процессе стационарных исследований. Описания растительности производились на пробных площадках размером 100 м2 (Шенников А.П., 1964). Участие отдельных видов в сложении каждого фитоценоза определялось на основе учета их обилия (по шкале Друде). Жизненные формы растений выделялись на основе классификации, предложенной А.П. Шенниковым (1950), В.М. Катанской (1981). При определении видовой принадлежности растений были использованы следующие источники: «Флора Средней полосы Европейской части СССР» (Маевский П.Ф., 1964), «Флора Европейской части СССР» под редакцией А. Федорова (1981), «Определитель растений Горьковской области» (Аверкиев Д.С., Аверкиев В.Д. 1985), «Конспект флоры Оренбургской области» (Рябинина З.Н., 1998). Номенклатура видов сверена по сводке С.К. Черепанова (1995).
Определение наличия и содержания биологически активных веществ проводили методами принятыми Всероссийским Институтом Лекарственных Растений (ВИЛР) и Институтом биохимии растений РАН (Биохимические методы анализа растений, 1960; Бандюкова, 1965; Государственная фармакопея СССР., 1968; Гришина, Самойлова, 1971; Булатов, Калинин, 1976; Краснов и др., 1985; Методы биохимического исследования растений под ред. Ермакова, 1989).
Определение числа сапрофитных бактерий проводилось согласно «Унифицированным методам исследования качества вод» (1985). Лизоцимная и антилизоцимная активность бактерий определялась чашечным методом с использованием принципа отсроченного антагонизма по отношению к тест-культуре Micrococcus lysodeikticus (Бухарин О.В. и др., 1984). При определении уровня лизоцимной активности водных макрофитов использовался метод фотометрического (турбидиметрического) определения лизоцимной активности (Каграманова К.А., Ермольева З.В., 1966) с использованием суспензий живых и обработанных хлороформом (Бухарин О.В. и др., 1997) клеток индикаторной культуры Micrococcus lysodeikticus (штамм № 2665 ГНИИСКМБП им. Л.А. Тарасевича).
Идентификацию выделенных штаммов микроорганизмов проводили до вида, а в некоторых случаях до рода, общепринятыми методами на основании морфологических, тинкториальных, культуральных и биохимических свойств. В качестве руководства по идентификации бактерий использовался Определитель бактерий Берджи (Holt I.G et al., 1997). При определении биохимических свойств выделенных микроорганизмов использовали коммерческие тест-системы фирмы LACHEMA (Чехия).
Математическую обработку полученных результатов проводили с помощью стандартных пакетов программ Statistica и Advanced Grapher 2.08, MS Excel 2000. РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЯ.
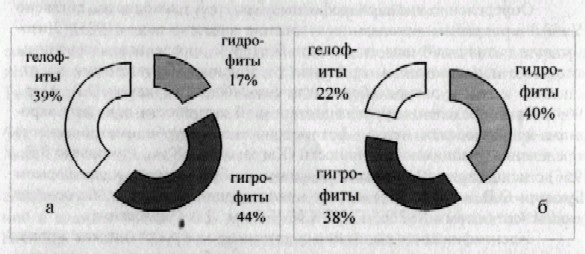
Изучение биологического разнообразия природных макро-фитно-бактериальных сообществ. Флора водных и прибрежно-водных сообществ лотических (р. Урал) и лентических (оз. Коровье, оз. Микутка) экосистем представлена 3 отделами: Bryophyta, Polypodiophyta, Magno-liophyta. Наиболее малочисленными были отделы Bryophyta, Polypodiophyta, каждый из которых представлен лишь одним семейством и составляет по 4% всей флоры сообществ. Отдел Magnoliophyta представлен 23 семействами, что составляет 92% флоры. В целом водная и прибрежно-водная растительность лотических и лентических экосистем представлена 38 видами, относящимся к 25 семействам и 28 родам. Из них 20 видов встречается исключительно в озерных биоценозах*, 6 видов - являются речными**, а остальные 12 видов являются общими***. При сравнении состава флоры р. Урал и двух озер в районе г. Оренбурга можно заметить ее достаточное своеобразие. Коэффициент сходства Жаккара лотических и ленточных экосистем равен 86% (полная флора). В ходе работы была проведена экологическая классификация растений исследуемых водоемов (рис. 1 а, б).
Рисунок 1 - Экологический спектр флоры водных и прибрежно-водных сообществ (а - р. Урал; б - озера)
Наибольшее количество исследуемых видов р. Урал относятся к гигрофитам, что составляет 44% от всей выявленной флоры (8 видов): Carex acuta*** С. riparia***, Rubus caesius*** , Galiwn palustre***, Lysimachia vulgaris*** , Myosotis palmtris*** , Veronica anagallis-aquatica ***, Bidens tripartite***. Гелофиты представлены 39% (7 видов): Typha angustifolia***, Т. latifolia **, Sparganium microcarpum **, Alisma plantago-aquatica***, Sagittaria sagittifolia ***, Scirpus lacustris ***, Butomm umbellatus **). Среди гидрофитов встречаются, только гидрофиты погруженные в воду, укореняющиеся - 17% (Potamogeton crispus**, P. perfoliatus***, Najas major**).
На озерах водная и прибрежно-водная растительность широко представлена гидрофитами, на долю которых приходится 40% от всей флоры (13 видов): гидрофиты укореняющиеся, с плавающими на поверхности листьями - 9 % (Persicaria amphibian*, Nuphar luted*, Hydrocharis morus-ranae*). Гидрофиты погруженные в воду, укореняющиеся - 13% (Potamogeton lucens* , P. perfoliatus *, Riccia fluitans* , Zannichellia palustris*). Гидрофиты не укореняющиеся, свободно плавающие на поверхности воды - 3% (Salvinia natans*). Гидрофиты погруженные в воду и не укореняющиеся - 16% (Lemna minir*, L. trisuica* , Elodea Canadensis* , Ceratophyllum demersum* , C. submersum*). На долю гигрофитов приходится 12 видов исследуемой флоры, что составляет 38% (12 видов). Среди гигрофитов представлены: Carex acuta***, С. riparia***, A triplex littoralis*, Caltha palmtris* , Ranunculus repens* , Rubus caesius*** , Myriophyllum spicatum* , Lysimachia vulgaris*** , Myosotis palustris*** , Veronica anagallis-aquatica*** , Galium palustre* , Bidens tripartite***. Соответственно гелофиты представлены 7 видами (Typha angustifolia***, Alisma gramineum***, A. plantago-aquatica*, Sagittaria sagittifolia*, Scirpus lacustris***, Phragmites australis*, Ph. communis*), что составляет 22%.
Таким образом, флора лотических и лентических типов экосистем отличается разнообразием видов и экологическим спектром. Флора лентических экосистем (озер) представлена 32 видами, а флора лотических экосистем (река) лишь 17 видами. Основу экологического спектра лентических экосистем составляют гидрофиты (40%) и гигрофиты (38%), а лотических - гигрофиты (44%) и гелофиты (39%). Доминантными видами макрофитов лотических экосистем являются Potamogeton perfoliatus, Potamogeton crispus, Najas major; лентических экосистем - Salvinia natans, Ceratophyllum demersum, Hydrocharis morus-ranae.
Проводимые нами исследования бактериопланктона лотических и лентических экосистем показали, что общая численность бактерий и количество сапрофитной микрофлоры подчиняются одинаковым межгодовым и сезонным закономерностям. Так, в 2002 году ОЧБ на р. Урал колебалась в среднем от 1021 кл/мл до 6121 кл/мл, а на озерах от 8235 до 11020кл/мл. То есть разница между максимальными величинами ОЧБ в реке и озерах составляла - 1,8 раза, а между минимальными - 8,0 раз. В 2003 году разница максимальных величин ОЧБ в этих экосистемах была соответственно 1,8 раза, минимальных - 2,9 раза, в Урале она колебалась от 2900кл/мл до 13980 кл/мл, а в озерах от 8500 кл/мл до 25000 кл/мл. По всей вероятности, такое повышение численности бактерий в 2003 году связано с поступлением аллохтонной микрофлоры с водосборной площади, так как в этот год интенсивно выпадали осадки и уровень воды в водоемах был высоким (Гидрологический ежегодник, 2002, 2003). В сезонной динамике общей численности бактерий были отмечены сходные закономерности как в лотических, так и лентических экосистемах. В 2002 году как на р. Урал, так и на озерах четко фиксируются три максимума: май, июль, сентябрь. На р. Урал минимальное и максимальное ОЧБ на ст. Водозабор и Железнодорожный мост составило в мае - 4469 кл/мл и 4964 кл/мл; в июле -5565 кл/мл и 6121 кл/мл; в сентябре -4590 кл/мл и 5327 кл/мл. На озерах ОЧБ была примерно одинаковой и составила в мае на оз. Микутка 8235 КОЕ/мл, на оз. Коровье 8707 КОЕ/мл; в июле 10976 КОЕ/мл и 11020 КОЕ/мл соответственно; в сентябре 8974 и 10330 КОЕ/мл соответственно. В 2003 году отмечалось повышение общей численности бактерий по сравнению с 2002 годом, при этом отмечалось постепенное увеличение общего числа бактерий как в лентических, так и в лотических экосистемах с мая по октябрь. На р. Урал наименьшая ОЧБ в 2003 году наблюдалась на ст. Водозабор, а наибольшая на ст. Железнодорожный мост. В мае ОЧБ на ст. Водозабор составила 2900 кл/мл, а в сентябре 12300 кл/мл, а на ст. Железнодорожный мост в эти же месяцы общая численность бактерий колебалась от 3428 кл/мл до 13884 кл/мл. На озерах ОЧБ также увеличивалась с мая по октябрь и составила в мае на оз. Микутка 5438 кл/мл, на оз. Коровье 6112 кл/мл, а в октябре соответственно 25800 кл/мл и 25707 кл/мл.
Также отмечена разница в количестве сапрофитной микрофлоры лотических и лентических экосистем. В среднем количество сапрофитной микрофлоры было выше на озерах в 1,6-2,2 раза по сравнению с рекой. В 2002 году их количество изменялось в Урале от 365 КОЕ/мл до 3100 КОЕ/мл, а в озерах от 787 КОЕ/мл до 4780 КОЕ/мл. В 2003 от 232 КОЕ/мл до 7300 КОЕ/мл в р. Урал, и от 537 КОЕ/мл до 11387 КОЕ/мл в озерах. В сезонной динамике изменения ОЧБ и количестве сапрофитной микрофлоры были отмечены одинаковые закономерности как в лотических так и лентических экосистемах: в 2002 году как в реке, так и в озерах зафиксировано три пика повышения численности бактериопланктона (май, август, сентябрь), а в 2003 году шло постепенное увеличение бактерий с мая по сентябрь.
Изучение микробных сообществ лотических и лентических экосистем показало, что в микробных популяциях этих водоемов широко представлены как лизоцимные, так и антилизоцимные формы бактерий, численность которых варьировала в разных водоемах и по сезонам года. В р. Урал количество лизоцимактивных форм бактерий колебалось в среднем от 26+10 до 36+10 КОЕ/мл, что составило 2,8-2,9% от общего числа сапрофитов. В озерах лизоцимактивные формы составили в среднем 45+10 - 74+30 КОЕ/мл, это 2,6-2,9% от общего числа сапрофитов. Численность антилизоцимактивных форм бактерий в р. Урал по среднегодовым данным составила от 28+15 до 53+32 КОЕ/мл, это 2,5-3,2% от общего числа сапрофитов. На озерах численность антилизоцимактивных форм колебалась от 39+25 до 54+32 КОЕ/мл, что составляет 2,2-3,5% от общего числа сапрофитной микрофлоры. В ходе исследования было установлено, что изменение численности лизоцимактивных и антилизоцимактивных форм бактерий коррелировало с изменением численности сапрофитов в экосистемах. Так коэффициент корреляции в лотических и лентических экосистемах между сапрофитными и лизоцимактивными формами бактерий составил 0,95-0,96, а между сапрофитами и антилизо-цимактивными формами бактерий 0,19-0,98. Соотношение ЛА и АЛА форм бактерий (Л) характеризует состояние водного биоценоза, и позволяет выявлять ранние сдвиги в гомеостазе водных экосистем, а также может служить критерием оценки экологического состояния водоема (Соловых Г.Н., 1995). Анализ результатов оценки экологического состояния исследованных водоемов по соотношению ЛА и АЛА форм бактерий показал, что в среднем индекс Л на р. Урал был меньше единицы в июле, августе и сентябре и составил соответственно 0,83; 0,56; 0,70, что говорит об экологическом неблагополучии реки в этот период, в то время как в мае, июне и октябре экологическое состояние реки улучшилось, о чем свидетельствует тот факт, что индекс Л был больше единицы: 1,50; 1,35; 1,63. На озерах индекс Л был всегда выше единицы и составил в среднем 1,15-1,57, что свидетельствует об экологическом благополучии этих водоемов. Таким образом, анализ результатов оценки экологического состояния исследованных водоемов по соотношению ЛА и АЛА форм бактерий показал, что наиболее благоприятная ситуация отмечалась на озерах, в то время как на р. Урал оно постоянно менялось.
Одним из эффективных путей изучения морфологического разнообразия микрофлоры, позволяющим судить о характере бактериоценозов водоемов, является использование стекол обрастания (Родина А.Г., 1966). Просмотр стекол обрастания показал, что независимо от сезона года, гидрохимических и гидробиологических условий преобладающее место в бактериопланктоне Урала и озер занимали кокки, их количество на разных разрезах составляло от 49 до 99% от общего числа бактерий. Далее следовали палочки как крупные, так и мелкие, численность которых обычно составляла 14-48% от общего количества бактерий. Количество нитевидных клеток не превышало 0,5-35% от общего числа бактерий.
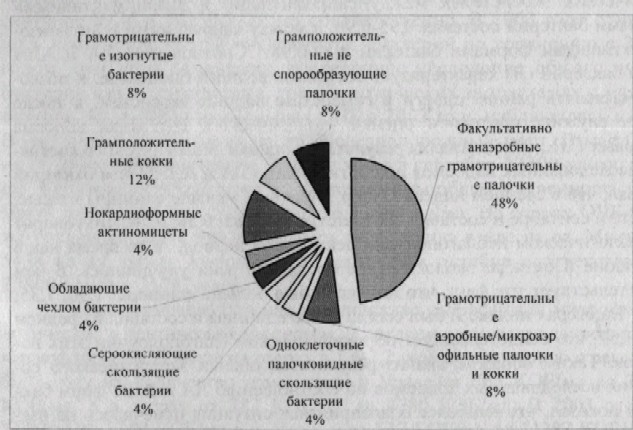
Проведенная идентификация выделенных штаммов микроорганизмов по морфологическим, тинкториальным, культуральным, биохимическим свойствам не выявила видовых различий бактериопланктона в лотических и лентических экосистемах. Согласно классификации Д. Берджи исследуемые виды относятся к 9 группам (рис. 2), которые выделены в 26 родов; определено 16 видов относящихся к различным таксономическим группам и семействам. В весенне-летний период в бактери-опланктоне лотических и лентических экосистем постоянно присутствовали микроорганизмы 4 родов (Enterobacter, Proteus, Providencia, Pseudomonas), а в осенний период - 3 родов (Ar throb act er, Cardiobacteriwn, Nocardia).
Рисунок 2 Группы основных категорий микрофлоры воды исследуемых водоемов по Д. Берджи (доля видов образующих данную группу категорий бактерий, %)
Таким образом, при исследовании общей численности бактерий и количества сапрофитной микрофлоры изученных водоемов выявлено превышение ОЧБ в озерах по сравнению с р. Урал в 1,8-8 раз, а количество сапрофитной микрофлоры в 1,5-2,2 раза. Отмечены межгодовые изменения численности бактерий, обусловленные колебанием уровня воды в водоемах; в сезонной динамике бактериопланктона установлены пики как общей численности бактерий, так и количестве сапрофитов, которые одинаковы в лотических и лентических экосистемах. Процентное соотношение численности ЛА и АЛА форм бактерий от числа сапрофитов одинаково как в лотических, так и лентических экосистемах; количество ЛА и АЛА форм лотических и лентических экосистем находятся в корреляции с численностью сапрофитов; сезонная динамика лизоцимактивных и антилизоцимактивных форм бактерий лотических и лентических экосистем различна: в лотических экосистемах в мае, июне и октябре преобладающей группой являются лизоцимактивные формы бактерий, а в июле, августе и сентябре антилизоцимактивные формы, а в лентических экосистемах преобладающей группой являются лизоцимактивные формы бактерий. Таксономическое определение микрофлоры лотических и лентических экосистем не выявило различий. Согласно классификации Берджи выявленные микроорганизмы были отнесены к 9 группам основных категорий, из них наиболее многочисленными являются факультативно анаэробные грамотрицательные палочки (48% от общего числа бактерий). Характеристика биологических свойств макрофитов (лизоцимная активность, содержание флавоноидов, аскорбиновой кислоты). В ряде работ показано наличие лизоцима у растений (Ермольева З.В., 1938; Кравченко Н.А. и др., 1965, 1972; Страйер Л., 1984; Лахтин В.М. и др., 1995; Smith, 1955), но нет данных о лизоцимной активности водных макрофитов. Поэтому на следующем этапе была поставлена задача оценить лизоцимную активность у доминантных видов водных макрофитов исследуемых водоемов. Проведенные исследования показали, что экстракты тканей доминантных видов макрофитов лотических и лентических экосистем проявляют лизоцимподобную активность в отношении суспензии как живых, так и убитых хлороформом клеток тест-культуры М. lysodeikticus (табл. 1), при этом она была ниже активности яичного лизоцима при концентрации белка 1 мкг/мл.
Таблица 1. Сравнительная характеристика лизоцимной активности экстрактов макрофитов исследуемых водоемов
виды растений
относительная активность (ед. а./мл)
удельная литическая активность (ед.а./мг белка)
Potamogeton perfoliatm (вегетативная часть)
1,6±0,2
0,31±0,05
Potamogeton crispus (вегетативная часть)
1,8±0,3
0,32±0,11
Najas major (вегетативная часть)
0,6±0,1
0,18±0,1
Hydrocharis morus-ranae (вегетативная часть)
2,0±0,11
0,52±0,02
Ceratophyllitm demersum (вегетативная часть)
1,2±0,2
0,48±0,3
Salvinia natans (вегетативная часть)
1,4±0,3
0,34±0,11
Наиболее высокое значение мурамидазной активности наблюдалось в вегетативной чисти Hydrocharis morus-ranae и составило 0,52+0,02 ед.а./мг белка, а наиболее низкое в вегетативной части у Najas major 0,18+0,1 ед.а./мг белка, что в 14-41 раз ниже активности яичного лизо-цима, которая равна 7,33 ед.а./мг белка. Наблюдаемое литическое действие экстрактов растений в отношении убитых клеток М. lysodeikticus может быть обусловлено продукцией лизоцима и, таким образом, может рассматриваться как проявление мурамидазной активности.
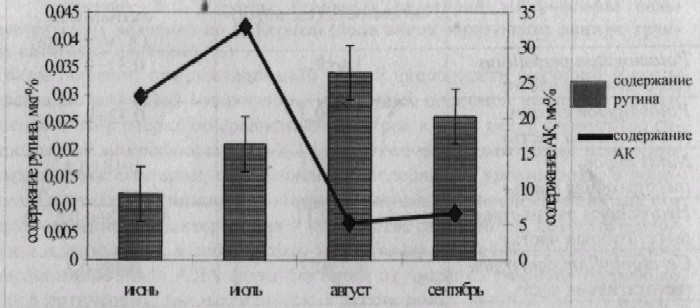
Проведенный анализ литературных данных свидетельствует об огромном значении БАВ в биологических процессах, протекающих в растительных и животных организмах, а потому в ходе нашей работы представлялось актуальным провести фитохимические исследования по оценке общего содержания флавоноидов, рутина и аскорбиновой кислоты, оказывающих влияние на биологические процессы в растительных организмах. Для исследования были взяты доминантные виды водных макрофитов лотических (Potamogeton perfoliatus, Potamogeton crispus, Najas major} и лентических экосистем (Hydrocharis morus-ranae, Ceratophyllurn demersum, Salvinia natans, Nuphar luteum). Наиболее высокое содержание флавоноидов зафиксировано у Potamogeton crispus, Hydrocharis morus-ranae, Salvinia natans, а самое низкое у Najas major и Nuphar luteum. Содержание рутина и аскорбиновой кислоты у доминантных видов водных макрофитов было различным. Отмечено четкое увеличение содержания рутина в период бутонизации и цветения макрофитов. Например, у Hydrocharis morus-ranae в июне и июле соответственно количество рутина составляло 0,012 мкг% и 0,021 мкг%, а в августе (период цветения) -0,034мкг% (рис. 3).
Рисунок 3 - Содержание рутина и АК у Hydrocharis morus-ranae в разные периоды вегетации
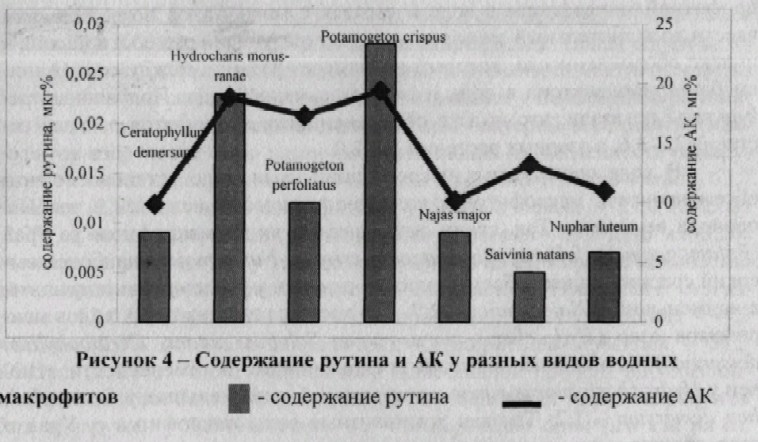
Различным было и содержание аскорбиновой кислоты у одного и того же вида в разные периоды вегетации (рис. 3). В ходе работы установлено максимальное содержание рутина у Hydrocharis morns-ranae (0,023+0,002 мкг%), Potamogeton crispm (0,028+0,001мкг %), минимальное у Salvinia natans (0,004+0,001 мкг%) (рис. 4). Исследование содержания аскорбиновой кислоты в зеленной массе водных макрофитов выявило отличия по этому показателю у разных видов: максимальное содержание аскорбиновой кислоты отмечено у Potamogeton crispm (19,4+0,2 мг%), а минимальное у Ceratophyllum demersum (9,9+0,2 мг%) (рис. 4).
Оценка макрофитно-бактериального состояния в водном биоценозе. Длительное существование организмов в рамках одной экосистемы возможно лишь в том случае, когда их компоненты и элементы дополняют друг друга и соответственно приспособлены друг к другу (Реймерс Т., 1994). Многочисленность и разнонаправленность действия биологических веществ, продуцируемых бактериями и макрофитами, приводит к вопросу о возможности существования комплекса макрофит-бактерии как некой системы объектов и взаимодействий, характеризующихся резервом устойчивости, обеспечивающим функционирование и развитие ее в условиях действия факторов среды. В связи с этим не вызывает сомнения актуальность следующего этапа исследования, затрагивающего вопросы механизмов взаимодействия про- и эукариот в системе «бактерии-макрофиты». Поэтому была поставлена задача, выяснить эколого-физиологические связи между микроорганизмами и макрофитами в природных водоемах.
Анализ содержания микрофлоры в биотопе открытых водоемов и микробных ассоциаций макрофитов показал, что в пробах воды общее число микроорганизмов было ниже, чем в смывах с водных макрофитов. Так, на р. Урал количество сапрофитной микрофлоры в воде в среднем составило 3006 КОЕ/мл, а в смывах с доминантных видов макрофитов от 8227 КОЕ/мл до 11920 КОЕ/мл. На озерах количество сапрофитной микрофлоры составило в среднем 4467 КОЕ/мл, а в смывах с макрофитов от 5138 КОЕ/мл до 6258 КОЕ/мл. Выявленные различия в количестве сапрофитной микрофлоры в воде и смывах с макрофитов позволили нам ввести дополнительный показатель, характеризующий это соотношение - индекс обсемененности, который показывает разницу между содержанием бактериопланктона в воде и смывах с макрофитов. Полученные результаты показали, что индекс обсемененности макрофитов р. Урал составил 2,2-3,6, а озерных растений 1,5-2,0.
В ходе проведенных исследований нами было установлено, что обсемененность макрофитов в воде неодинакова и меняется в течение периода вегетации. Так, среди доминантных видов макрофитов р. Урал (Potamogeton perfoliatm, Potamogeton crispus, Najas major) наиболее высокий средний индекс обсемененности отмечен у Potamogeton crispus - 4, а наименьший у Najas major - 2,7. Что касается доминантных видов макрофитов озер (Hydrocharis morus-ranae, Salvinia natans, Ceratophyllum demersum), то наиболее высокий средний индекс обсемененности отмечен у Hydrocharis morus-ranae и составил 1,4, а наименьших у Ceratophyllum demersum - 1,2. JTaK как доминантные виды макрофитов р. Урал и озер отличны, то не представлялось возможным сравнивать их индекс обсемененности между собой, однако заметно, что у макрофитов р. Урал он выше, чем у макрофитов озер. Следует также отметить, что обсемененность водных макрофитов изменяется по месяцам в течении периода вегетации.
Анализ состава микробиоценоза по их функциональной активности показал, что бактериальные популяции в воде представлены как лизоцимактивными, так и антилизоцимактивными формами бактерий, однако среди бактерий ассоциантов макрофитов были представлены только антилизоцимактивные формы. За весь период исследований в реке Урал нами не обнаружено в смывах с растений лизоцимактивных форм бактерий, в тоже время антилизоцимактивные формы встречались. Так на станции «Водозабор» количество антилизоцимактивных форм бактерий ассоциантов макрофитов составило в среднем у Potamogeton perfoliatm -114,5±20 КОЕ/мл, у Potamogeton crispus - 116,1+20 КОЕ/мл, у Najas таjor 86,9+2,0 КОЕ/мл, что соответственно составляет 1,6+0,9%, 1,6+0,7%, 2,8+1,1% от числа сапрофитных бактерий, ассоциантов мак-рофитов. Наибольший процент антилизоцимактивных форм бактерий от числа сапрофитов на станции «Водозабор» наблюдался у Najas major и составил 2,8+1,1%, а наименьший у Potamogeton perfoliatus - 1,6+0,9%. Количество антилизоцимактивных форм на станциях Автодорожный и Железнодорожный мост было примерно одинаковым, но отличалось от такового на станции Водозабор. Так, количество антилизоцимактивных форм у Potamogeton perfoliatm составило на этих станциях соответственно 66,7 и 75,5 КОЕ/мл (0,9+0,4% от числа сапрофитов); у Potamogeton crispus 69,4 и 77,5 КОЕ/мл (0,7+0,1% от числа сапрофитов); у Najas major 64,0 и 72,3 КОЕ/мл (1,2+0,2 % от числа сапрофитов). Таким образом, на всех станциях р. Урал наибольший процент антилизоцимактивных форм бактерий наблюдался у Najas major, а наименьшее у Potamogeton perfoliatus Количество антилизоцимактивных форм бактерий ассоциантов макрофитов озер также было различно, наибольшая их численность обнаружена в смывах с макрофитов оз. Микутка: Hydrocharis morus-ranae - 67,7 КОЕ/мл (2,8% от числа сапрофитов), у Salvinia natans - 59,0 КОЕ/мл (3,2% от числа сапрофитов), Ceratophyllum demersum - 65,3 КОЕ/мл (3,2% от числа сапрофитов). А у этих же макрофитов на оз. Коровье количество антилизоцимактивных форм бактерий составило у Hydrocharis morus-ranae - 52,0 КОЕ/мл (2,3% от числа сапрофитов), у Salvinia natans — 44,3 КОЕ/мл (2,8% от числа сапрофитов), Ceratophyllum demersum - 51,2 КОЕ/мл (2,8% от числа сапрофитов). Наибольший процент антилизоцимактивных форм бактерий ассоциантов макрофитов от числа сапрофитов был обнаружен у Salvinia natans и у Ceratophyllum demersum как на оз. Микутка, так и на оз. Коровье.
Таким образом, в результате проведенного исследования установлено, что в лотических и лентических экосистемах по функциональной активности бактерии-ассоцианты макрофитов представлены лишь анти-лизоцимными формами, их количество в смывах у различных видов макрофитов отлично и в среднем составило: у Potamogeton perfoliatus -115+20 КОЕ/мл, у Potamogeton crispus - 116+25 КОЕ/мл, у Najas major -87+10 КОЕ/мл, у Hydrocharis morus-ranae - 224+26 КОЕ/мл, у Salvinia natans - 168+20 КОЕ/мл и у Ceratophyllum demersum - 220+24 КОЕ/мл (табл. 2).
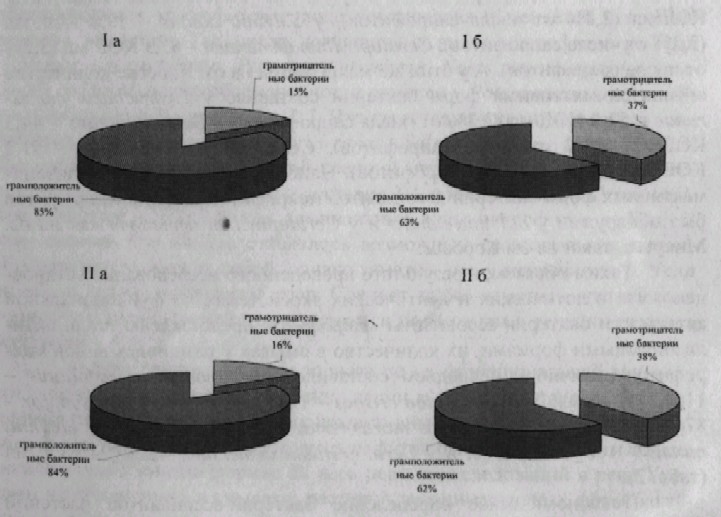
Таксономическое определение бактерий-ассоциантов растений показало, что видовой состав бактерий ассоциантов лотических и лентических экосистем не отличается особым разнообразием. Среди них высевались микроорганизмы, принадлежащие к родам Enterobacter, Pseudomonas Nocardia, Alcaligenes, Arthrobacter, Bacillus, Neisseria. В весенне-летний период в смывах с макрофитов отмечены представители родов Enterobacler, Providencia, а в осенний период нокардиоформные актино-мицеты, представители родов Pseudomonas и Enterobacter. При анализе ассоциаций бактерий в воде и смывах с растений были обнаружены следующие закономерности: в воде на всех станциях преобладающей морфологической группой бактерий были грамположительные кокки, а в смывах с растений, то есть в ассоциациях макрофиты-бактерии их процент заметно снижался и возрастало количество грамотрицательных кокков. Так в смывах с водных макрофитов р. Урал в целом процент грамотрицательных бактерий составил 36,3%, а в воде - 15%, то есть грамотрицательных бактерий в смывах с макрофитов было в 2,4 раза больше чем в воде (р<0,05) (рис. 6, I). Аналогичные закономерности были выявлены при исследованиях смывов с озерных растений: доля грамотрицательных бактерий в смывах - 38%, а в воде - 16%, то есть разница в 2,4 раза (р«М)5) (рис, 6, II).
Рисунок 6 - Морфологические группы бактерий (I - в реке Урал; II - в озерах; а - в воде; б - в смывах с растений)
Механизмы взаимоотношений в системе макрофиты-бактерии. Характер взаимодействия бактерий с высшими растениями неоднозначен: в одних случаях бактерии играют роль фитопатогенов, вызывающих заболевания и гибель растений, в других - симбионтов, которые защищают растения от патогенных микроорганизмов (в частности в ризосфере) (Lewin D., 1974; Тихонович И.А., Покровский Н.А., 1993). В качестве механизмов взаимодействия микрофлоры и гидробионтов может рассматриваться динамическая система, в которой с одной стороны выступают факторы естественной защиты, продуцируемые эукариотической клеткой (лизоцим), а с другой - направленная деградация этих факторов прокариотами (антилизоцимная).
Выявлена прямая корреляционная зависимость между общей численностью бактерий и уровнем лизоцимной активности макрофитов (г=0,83), а также между численностью антилизоцимактивных форм бактерий и уровнем лизоцимной активности (г=0,95) (табл.2). Так, среди доминантных видов макрофитов наиболее высокая лизоцимная активность, общая численность бактерий и количество антилизоцимактивных форм бактерий наблюдалась у Hydrocharis morus-ranae и составили соответственно 0,52+0,02 ед.а./мг белка, 24143±862 кл/мл и 224+20 КОЕ/мл, а самые низкие значения обнаружены у Najas major, которые составили 0,18+0,10 ед.а./мг белка, 13860+427 кл/мл и 87+20 КОЕ/мл.
Таблица 2. Уровень лизоцимной активности макрофитов и количество на них бактерий-ассоциантов
Виды
Удельная литическая активность (ед.а./мг белка)
Общая численность бактерий (кл/мл)
АЛА-формы бактерий (КОЕ/мл)
Potamogeton perfoliatus
0,31±0,05
19775±647
115±20
Potamogeton crispm
0,32±0,11
20690±842
116±25
Najas major
0,18±0,10
13860±427
87±10
Hydrocharis morus-ranae
0,52±0,02
24143±862
224±26
Ceratophyllum demersum
0,48±0,30
21272±774
220±24
Salvinia natans
0,34±0,11
22988±792
168±20
Учитывая роль лизоцима как защитного фактора беспозвоночных животных (Купер Э., 1980; Ломакина М.С., 1990), а также выявленную зависимость между численностью антилизоцимактивных форм бактерий-ассоциантов и уровнем лизоцимной активности макрофитов, нами разработан «Способ биологического обеззараживания непроточных водоемов от патогенной и условно-патогенной микрофлоры» (патент РФ на изобретение № 2193531 от 27 ноября 2002 года), позволяющий путем отбора и интродукции в биологические пруды гидробионтов - продуцентов лизоцима обеспечить обеззараживание воды от патогенной и условно-патогенной микрофлоры.
Богатство видового разнообразия макрофитов и бактерий с одной стороны, и высокий репродуктивный потенциал и метаболическая пластичность самих бактерий с другой, вероятно обуславливают многообразие форм взаимодействий популяции бактерий и растений. Становление и развитие подобных форм взаимодействий обеспечивается явлениями межвидового антагонизма и синергизма, обусловленных продукцией макрофитами и бактериями разного рода веществ: БАВ (флавоноиды, фитонциды и др.), антибиотики, лизоцим. Поэтому цель следующего этапа исследования была: оценить участие некоторых биологически активных веществ исследуемых макрофитов в формировании физиолого-экологических взаимоотношений в связке «макрофиты-бактерии» в природных водоемах (табл. 3).
Таблица 3. Количество микроорганизмов-ассоциантов и содержания БАВ у макрофитов
виды макрофитов
количество микроорганизмов-ассоциантов макрофитов (кл/мл)
флавоноиды
содержание аскорбиновой кислоты, (мг%)
проба Синода
содержание рутина (мкг%)
Potamogeton perfoliatus
19775±647
++
0,012±0,001
17,3±0,1
Potamogeton crispus
20690±842
+++
0.028±0.001
19,4±0,2
Najas major
13860±427
+
0,009±0,001
10,2±0,1
Hydrocharis morus-ranae
24143±862
+++
0,023±0.002
18.89±0,1
Ceratophyllum demersum
21272±774
++
0,009±0,001
9.9±0,2
Salvinia natans
22988±792
+++
0,004±0.001
13,2±0,2
Была установлена корреляционная зависимость между количеством общего содержания флавоноидов и общей численностью бактерий -ассоциантов этих макрофитов. Коэффициент корреляции этих параметров составил 0,9. Следовательно, флавоноиды макрофитов являются фитогенным фактором, который влияет на рост бактерий.
Обобщая полученный фактический материал, следует отметить, что биоценозы лотических и лентических водоемов характеризуются биологическим разнообразием макрофитно-бактериальных сообществ, уровень которого зависит от типа экосистемы. Полученные данные позволили обосновать участие флавоноидов водных растений и системы «лизоцим макрофитов - антилизоцим бактерий» в формировании макрофитно-бактериальных ассоциаций гидробиоценозов.
ВЫВОДЫ
1. Флора водных и прибрежно-водных сообществ макрофитов лотических и лентических экосистем отличается видовым разнообразием, представлена 38 видами, относящимися к 25 семействам и включающим 3 отдела (Bryophyta, Polypodiophyla, Magnoliophytd). Основу экологического спектра лентических экосистем составляют гидрофиты (40%) и гигрофиты (38%), а лотических - гигрофиты (44%) и гелофиты (39%). Доля общих видов для этих экосистем составляет 86%. Доминантными макрофитами лотических экосистем являются Potamogeton perfoliatus, Potamogeton crispus, Najas major; а лентических - Salvinia natans, Ceratophyllum demersum, Hydrocharis morus-ranae.
2. Выявлено сходство видовых составов бактериопланктона лотических и лентических экосистем, которые представлены микроорганизмами из 26 родов, относящихся к 9 таксономическим группам. Отмечена сезонная динамика видового разнообразия бактериопланктона: в весенне-летний период в нем постоянно присутствовали микроорганизмы 4 родов (Entervbacter, Proteus, Pseudomonas, Providencia), а в осенний период - 3 родов (Arthrobacter, Cardiobacterium, Nocardia).
3. В структуре бактериопланктона лотических и лентических биоценозов преобладали кокковые (49-99%) и палочковидные (14-48%) микроорганизмы над нитевидными формами бактерий (0,5-35%). В исследуемых водоемах микрофлора была представлена как лизоцимактивными, так и антилизоцимактивными формами, соотношение которых зависело от типа экосистемы и было подвержено сезонным колебаниям. В лентических экосистемах численность антилизоцимактивных форм бактерий была ниже количества лизоцимактивных форм (2,7+0,1% от общего числа бактерий против 2,9+0,1%), тогда как в лотических бактериоценозах их доля в мае, июне, октябре была незначительной (0,16%; 0,11%; 0,14% от общего числа бактерий), но возрастала с июля по сентябрь (1,46%-0,82%) на фоне уменьшения частоты встречаемости лизоцимактивных форм бактерий (1,01 %-0,10%).
4. У водных макрофитов обнаружены флавоноиды, в том числе рутин, и аскорбиновая кислота, содержание которых зависело от их видовой принадлежности: максимальное количество флавоноидов отмечено у Hydrocharis morus-ranae, Potamogeton crispus, а минимальное у Najas major, максимальное содержание рутина выявлено у Hydrocharis morns-ranae (0,023+0,002 мкг%), минимальное у Salvinia natans (0,004+0,001 мкг%); наибольшее количество аскорбиновой кислоты у Potamogeton crispus (19,4+0,2мг%), наименьшее - у Ceratophyllum demersum (9,9+0,2 мг%). Содержание данных биологически активных веществ изменяется в зависимости от периода вегетации макрофитов. Экстракты тканей макрофитов обладали лизоцимной активностью, уровень которой варьировал у разных видов: среди доминантных макрофитов наиболее высокий ее уровень обнаружен у Hydrocharis morus-ranae, а самый низкий - у Najas major.
5. Выявлено превышение в 1,2-5,1 раза численности бактерий связанных с поверхностью водных растений, над количеством бактерио-планктона в воде. Показана зависимость степени микробной обсеменен-ности доминантных видов макрофитов от их видовой и экотопной принадлежности и периода вегетации. Обнаружены виды макрофитов с максимальной (Hydrocharis тогш-ranae, Salvinia natans) и минимальной (Najas major) бактериальной колонизацией. Полученные данные свидетельствуют о формировании в водных биоценозах лотических и лентических экосистем относительно стабильных макрофитно-бактериальных ассоциаций.
6. Установлены прямые корреляционные связи между численностью бактерий-ассоциантов макрофитов и содержанием в растениях флавоноидов и аскорбиновой кислоты, между уровнем лизоцимной активности макрофитов и количеством антилизоцимактивных форм бактерий-ассоциантов, что свидетельствует об участии изученных биологически активных веществ и системы «лизоцим макрофитов - антилизоцим бактерий» в формировании эколого-функциональных связей в макрофитно-бактериальных сообществах водных биоценозов. Полученные данные легли в основу разработки «Способа биологического обеззараживания непроточных водоемов от патогенной и условно-патогенной микрофлоры» (патент РФ на изобретение № 2193531 от 27 ноября 2002 года).
Список работ, опубликованных по теме диссертации
1. Соловых Г.Н. Лизоцим гидробионтов и его роль в способности водоемов к процессу самоочищения / Г.Н. Соловых, Е.М. Нефедова, Г.М. Устинова, Е.К. Раимова, Е.А. Рябцева // Гигиена и санитария - 2002.-№5.-С. 24-26.
2. Ушакова Е.И. Лизоцимная чувствительность родококков как фактор формирующий их микробиоценоз / Е.И. Ушакова, Г.М. Устинова // 6-ая Пущинская школа-конференция молодых ученых «Биология - наука XXI века»: тез. докл.-Пущино, 2002. том 3.-С.69.
3. Устинова Г.М. К вопросу о взаимодействии высших водных растений с бактериопланктоном / Г.М. Устинова, Л.Г. Фабарисова // Современные проблемы экологии, микробиологии и иммунологии: сб. тр.- Пермь.-2002.-С. 25-26.
4. Устинова Г.М. Роль высших водных растений в самоочищении водоема / Г.М. Устинова, Л.Г. Фабарисова // II международная конференция «Биоразнообразие и биоресурсы Урала и сопредельных территорий»: тез. докл.-Оренбург, 2002.-С. 87-88.
5. Рябцева Е.А. К вопросу о разнообразии высших водных растений и сообществ простейших в водоемах города Оренбурга / Е.А. Рябцева, Г.М. Устинова // Региональная научно-практическая конференция молодых ученых и специалистов: тез. докл.-Оренбург, 2003.-С. 170-172.
6. Соловых Г.Н. Определение Р-витаминной активности у водных макрофитов / Г.Н. Соловых, Г.М. Устинова, Е.А. Рябцева // Межрегиональная конференция биохимиков Урала, Западной Сибири и Поволжья «Биохимия: от исследования молекулярных механизмов - до внедрения в клиническую практику и производство»: тез. докл.-Оренбург,2003.-С. 111-114.
7. Устинова Г.М. Макрофиты реки Урал и озер в районе города Оренбурга / Г.М. Устинова // Региональная научно-практическая конференция молодых ученых и специалистов: тез. докл.-Оренбург, 2004.-С. 27-28.
8. Устинова Г.М. Оценка макрофитно-бактериального состояния в водном биоценозе лотических и лентических экосистем / Г.М. Устинова, Г.Н. Соловых // II Всероссийская конференция «Экосистемы малых рек: биоразнообразие, экология, охрана»: тез. докл.- Борок, 2004. - С. 86-87.
9. Нефедова Е.М. Сравнительный анализ численности бактериопланктона лотических и лентических экосистем в районе города Оренбурга / Е.М. Нефедова, Л.Г. Фабарисова, Г.М. Устинова // II Всероссийская конференция «Экосистемы малых рек: биоразнообразие, экология, охрана»: тез. докл.- Борок, 2004. - С. 65
10. Соловых Г.Н. Способ биологического обеззараживания непроточных водоемов от патогенной и условно-патогенной микрофлоры / Г.Н. Соловых, В.П. Коробов, И.В. Карнаухова, Е.М. Нефедова, Г.М. Устинова / Патент РФ на изобретение № 2193531 от 27 ноября 2002 года, Бюл. №33. -14с.
 Германское издательство LAP LAMBERT Academic Publishing (LAP LAMBERT Academic Publishing is a trademark of: OmniScriptum GmbH & Co. KG) предлагает книгу на русском языке "Исторические исследования" (ISBN: 978-3-659-63167-2, цена - 89.90 ЕВРО), автор - Тихомиров Андрей Евгеньевич, аннотация на обложке "Современные археологические исследования доказывают, что родиной индоевропейцев является район Южного Урала, где они сформировались в качестве единой языковой группы. Аирйанэм-Ваэджа (авест. «арийский простор») — мифическая прародина древних иранцев, ариев; согласно Видевдату (Вендидаду, первой книге Авесты, сборника священных книг древнеиранской религии, своего рода иранское продолжение Вед), именно с неё начинается список из 16 «наилучших» «местностей и областей», созданных Ахура Маздой для человечества. Эта страна описывается как бескрайняя равнина, через которую протекает прекрасная река Даитйя (Вахви-Датия) - Урал".
Германское издательство LAP LAMBERT Academic Publishing (LAP LAMBERT Academic Publishing is a trademark of: OmniScriptum GmbH & Co. KG) предлагает книгу на русском языке "Исторические исследования" (ISBN: 978-3-659-63167-2, цена - 89.90 ЕВРО), автор - Тихомиров Андрей Евгеньевич, аннотация на обложке "Современные археологические исследования доказывают, что родиной индоевропейцев является район Южного Урала, где они сформировались в качестве единой языковой группы. Аирйанэм-Ваэджа (авест. «арийский простор») — мифическая прародина древних иранцев, ариев; согласно Видевдату (Вендидаду, первой книге Авесты, сборника священных книг древнеиранской религии, своего рода иранское продолжение Вед), именно с неё начинается список из 16 «наилучших» «местностей и областей», созданных Ахура Маздой для человечества. Эта страна описывается как бескрайняя равнина, через которую протекает прекрасная река Даитйя (Вахви-Датия) - Урал".